


The fact that the simple and deterministic equation (1) can possess dynamical trajectories which look like some sort of random noise has disturbing practical implications. It means, for example, that apparently erratic fluctuations in the census data for an animal population need not necessarily betoken either the vagaries of an unpredictable environment or sampling errors: they may simply derive from a rigidly deterministic population growth relationship such as equation (1). This point is discussed more fully and carefully elsewhere 1.
Alternatively, it may be observed that in the chaotic regime arbitrarily close initial conditions can lead to trajectories which, after a sufficiently long time, diverge widely. This means that, even if we have a simple model in which all the parameters are determined exactly, long term prediction is nevertheless impossible. In a meteorological context, Lorenz 15 has called this general phenomenon the "butterfly effect": even if the atmosphere could be described by a deterministic model in which all parameters were known, the fluttering of a butterfly's wings could alter the initial conditions, and thus (in the chaotic regime) alter the long term prediction.
Fluid turbulence provides a classic example where, as a parameter (the Reynolds number) is tuned in a set of deterministic equations (the Navier-Stokes equations), the motion can undergo an abrupt transition from some stable configuration (for example, laminar flow) into an apparently stochastic, chaotic regime. Various models, based on the Navier-Stokes differential equations, have been proposed as mathematical metaphors for this process 15, 40, 41. In a recent review of the theory of turbulence, Martin 42 has observed that the one-dimensional difference equation (1) may be useful in this context. Compared with the earlier models 15, 40, 41, it has the disadvantage of being even more abstractly metaphorical, and the advantage of having a spectrum of dynamical behaviour which is more richly complicated yet more amenable to analytical investigation.
A more down-to-earth application is possible in the use of
equation (1) to fit data
1,
2,
3,
38,
39,
43
on biological populations
with discrete, non-overlapping generations, as is the case for
many temperate zone arthropods. Figure 6
shows the parameter values 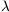 and
and 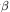 that are
estimated 39
for 24 natural populations
and 4 laboratory populations when equation (17) is fitted to the
available data. The figure also shows the theoretical stability
domains: a stable point; its stable harmonics (stable cycles of
period 2n); chaos. The natural populations tend to have stable
equilibrium point behaviour. The laboratory populations tend
to show oscillatory or chaotic behaviour; their behaviour may
be exaggeratedly nonlinear because of the absence, in a
laboratory setting, of many natural mortality factors. It is
perhaps suggestive that the most oscillatory natural population
(labelled A in Fig. 6) is the
Colorado potato beetle, whose
present relationship with its host plant lacks an evolutionary
pedigree. These remarks are only tentative, and must be treated
with caution for several reasons. Two of the main caveats are
that there are technical difficulties in selecting and reducing the
data, and that there are no single species populations in the
natural world: to obtain a one-dimensional difference equation
by replacing a population's interactions with its biological and
physical environment by passive parameters (such as
and
)
may do great violence to the reality.
that are
estimated 39
for 24 natural populations
and 4 laboratory populations when equation (17) is fitted to the
available data. The figure also shows the theoretical stability
domains: a stable point; its stable harmonics (stable cycles of
period 2n); chaos. The natural populations tend to have stable
equilibrium point behaviour. The laboratory populations tend
to show oscillatory or chaotic behaviour; their behaviour may
be exaggeratedly nonlinear because of the absence, in a
laboratory setting, of many natural mortality factors. It is
perhaps suggestive that the most oscillatory natural population
(labelled A in Fig. 6) is the
Colorado potato beetle, whose
present relationship with its host plant lacks an evolutionary
pedigree. These remarks are only tentative, and must be treated
with caution for several reasons. Two of the main caveats are
that there are technical difficulties in selecting and reducing the
data, and that there are no single species populations in the
natural world: to obtain a one-dimensional difference equation
by replacing a population's interactions with its biological and
physical environment by passive parameters (such as
and
)
may do great violence to the reality.
Some of the many other areas where these ideas have found applications were alluded to in the second section, above 5 - 11. One aim of this review article is to provoke applications in yet other fields.